В.Ю. Дмитриев
РАННЕМЕЛОВЫЕ ЭНТОМОКОМПЛЕКСЫ ЦЕНТРАЛЬНОЙ АЗИИ
Опубликовано онлайн 07 марта 2017 г.
ЦЕЛЬ РАБОТЫ
Работа предпринята для выяснения последовательности комплексов насекомых в нижнем мелу Центральной Азии. Стимулом явилась потребность разрешить противоречие привычной датировки Исяня как начала раннего мела с современной радиоизотопной датировкой как баррема-апта (Chang and oth., 2009; Gradstein and oth., 2012). Без прояснения этого вопроса оказалось затруднительно завершить начатое ранее в Лаборатории артропод обобщение по динамике разнообразия насекомых (Дмитриев и др., 2017, в печати)
ПОСТАНОВКА ЗАДАЧИ
В нижнем мелу Центральной Азии при современной изученности разрезов и энтомофауны отчетливо различаются два комплекса насекомых.
Более древний комплекс
характеризуется присутствием стрекозы Sinaeschnidia, более молодой – стрекозы Hemeroscopus (с единственным видом H. baissicus).
Sinaeschnidia найдена в китайских формациях Исянь, Лайян и Шахай (во всех трех S. heishankowensis), в тургинской свите Забайкалья и нижнецаганцабской
подсвите Монголии (Хутэл-Хара). Hemeroscopus baissicus обнаружен в Байсе (Забайкалье), катайской формации Лушанфен и
монгольских хурилтской, шартологойской , шинхудукской,
андахудукской, тэвшской и хухтыкской свитах, а также местонахождениях Хобур,
Баян-Дурах, Шине-Ус.
Имеются свиты и местонахождения, в которых характерные стрекозы не найдены. В некоторых случаях эти отложения могут быть отнесены к одному из указанных уровней по присутствию сопутствующих Sinaeschnidia и Hemeroscopus родов и видов (см. ниже).
Основной вопрос: формация Исянь древнее, моложе или одновременна с тургинским горизонтом Восточного Забайкалья ? Для ответа требуется выяснить последовательность комплексов более детально и желательно в более широком контексте.
Рассмотрены следующие комплексы:
1. Юрский (условное название) включает насекомых поздней юры (Каратау, Шар-Тэг, удинская свита, Хоутийн-Хотгор, Кемпендяй), частично средней юры (пачка Даохугоу, формации Юлоншан и Хайфангоу) и пограничных между ними отложений (Бахар).
2. Ундинодаинский комплекс составляют насекомые одноименного горизонта Восточного Забайкалья. Они происходят из глушковской, укурейской, тергенской, букукунской и чалунихинской свит (Атлас…, 2002).
Ундинодаинский горизонт относится к верхам юры, но здесь рассматривается отдельно. Названия “юра” и “юрский” далее в тексте везде подразумевают юру без ундинодаинских отложений. Юрский и ундинодаинский комплексы включены в состав анализируемой последовательности, так как в них есть много родов, найденных в отложениях с Sinaeschnidia и Hemeroscopus (в последних таких родов значительно меньше).
3. Тургинский комплекс это насекомые даинской, тургинской, гидаринской, устькарской, годымбойской и бянкинской свит Восточного Забайкалья (Атлас…, 2002).
В бянкинской свите собраны жуки Coptoclava (Абагайтуй), характерные для тургинских и более молодых отложений. Насекомые из других местонахождений бянкинской свиты равным образом встречаются в ундинодаинском и тургинском горизонтах и не влияют на характеристику их комплексов.
4. Исяньский комплекс происходит из многочисленных местонахождений в одноименной китайской формации.
5. Андахудукский комплекс объединяет монгольских насекомых андахудукской, шинхудукской, шарынгольской, тэвшской свит, а также уланаргалантской серии из района Бон-Цагана (см. Андахудукский горизонт в разделе “Биостратиграфия”).
6. Зазинский комплекс известен только из местонахождения Байса на р. Витим, единичные представители также из Романовки.
7. Хурилтский комплекс это насекомые хурилтской (главным образом в местонахождении Бон-Цаган) и шартологойской свит Цетральной Монголии (см. Зазинская свита и хурилтский горизонт в разделе “Биостратиграфия”).
Далее для краткости используются названия “андахудукский” и “хурилтский” горизонты, включающие перечисленные выше отложения.
Таким образом, здесь собраны вместе два восточнозабайкальских комплекса (ундинодаинский и тургинский), западнозабайкальский зазинский, центральномонгольские андахудукский и хурилтский и китайский исяньский. Стратиграфические соотношения известны для восточнозабайкальских и центральномонгольских горизонтов: ундинодаинский древнее тургинского, андахудукский древнее хурилтского. Также известно (см. выше), что Sinaeschnidia есть в тургинском и исяньском комплексах, а Hemeroscopus в зазинском, андахудукском и хурилтском. Хурилтский комплекс скорее всего является наиболее молодым.
Задача состоит в прояснении соотношений обоих комплексов с Sinaeschnidia и всех трех с Hemeroscopus.
МАТЕРИАЛ
Использовались следующие данные:
1. Списки насекомых, составленные М.М. Мостовским по коллекционным тетрадям Лаборатории с некоторыми его собственными дополнениями (сайт Лаборатории, palaeoentomolog.ru).
2. Там же более полные списки имеющихся в Лаборатории материалов по отрядам Plecoptera, Mecoptera и Diptera.
3. Списки А.Г. Пономаренко по мезозойским насекомым Монголии (там же)
4. Материалы монографий Лаборатории артропод по Сибири и Восточному Забайкалью (Позднемезозойские …, 1990; Калугина, Ковалев, 1985; Юрские насекомые…, 1985, Мезозойские…, 1993).
5.
Данные EDNA (edna.palass-hosting.org) и Paleobiology Database (paleodb.org)
по Китаю.
6. Дополнительные сведения получены непосредственно от сотрудников Лаборатории, а по ортоптерам местонахождения Семен - просмотром этикеток в коллекции.
Все сведения представлены в основной таблице. В ней серым цветом выделены роды, найденные более, чем в одном местонахождении (не выделены роды, встреченные только в юре); желтым цветом показано то же самое для видов. Положение некоторых родов в составе указанных для них семейств нуждается в дополнительной проверке. Пометка (cf.) в столбце Subgenus относится к роду, а в столбце Note – к виду.
Материал неполон и неоднороден. Монгольские коллекции изучены только частично. Сведения по Китаю собраны в указанных базах данных из публикаций разных лет. Обе базы данных неполны. Только около одной трети материалов каждой из них совпадает с данными другой базы. Только в американской базе данных приводятся (частично ?) сведения о результатах ревизий. Весь материал нуждается в более полном изучении и ревизии. Без этого трудно рассчитывать на получение вполне обоснованных выводов. Но вследствие ограниченного времени для данной работы не преследовалась цель исчерпывающего сбора данных. Предполагается, что перечисленные выше материалы достаточны для выяснения общей последовательности энтомокомплексов.
МЕТОДИКА
Использовались роды и виды, встреченные более, чем в одном комплексе. Под “комплексом” далее подразумеваются все только такие роды и виды.
Исключены роды и виды, не имеющие стратиграфического значения для меловых отложений, то есть известные только в юре, а также в юре или ундинодаинском горизонте и проходящие до хурилтского горизонта. Кроме того, исключены все индузироды и Coptoclava, встречающаяся на всех уровнях нижнемеловых отложений.
Сведения о распространении родов и видов приведены в таблице основных комплексов с семью столбцами. Таблица содержит 277 строк (239 родов, 64 вида, 26 родов найдены там же, где и единственный относящийся к ним вид). Ниже, в разделе “Побочные результаты” использованы сведения о ещё нескольких родах и видах, не обнаруженных в составе основных комплексов. В таблице использованы следующие обозначения основных комплексов: Jura - юрский, Unda - ундинодаинский, Turg - тургинский, Yixi - исяньский, Anda - андахудукский, Zaza - зазинский, Khur - хурилтский.
Основные комплексы сравнивались с помощью простого количественного показателя. Для каждых двух комплексов определялась доля общих родов в числе родов, составляющих меньший комплекс, характеризующая близость меньшего комплекса к большему. Если в составе комплекса имеются предположительно определенные роды (cf.), при подсчете они учитывались как 1/2.
Во втором варианте оценки использовались также данные о видах. Правила подсчета следующие:
общий род – 1,
то же с предположительным определением – 1/2,
общий вид – 2,
то же с предположительным определением – 1.
Вес 2 для общих видов подразумевает осторожное отношение к стратиграфическому значению видов при отсутствии данных об их точном стратиграфическом положении в ряде хорошо изученных разрезов. Возможно, общие виды более значимы. Поэтому использовался третий вариант с правилами подсчета:
общий род – 1,
то же с предположительным определением – 1/2,
общий вид – 5,
то же с предположительным определением – 2.
Суммы, используемые во всех трех вариантах, далее называются суммами баллов. Во втором и третьем вариантах суммы баллов использовались так же, как в первом.
Результаты отображены на гистограммах, в которых столбцы ранжированы по убыванию. Обозначения комплексов на гистограммах те же, что в таблице основных комплексов. На гистограммах при названиях комплексов всегда указана их общая сумма баллов, чтобы обозначить, какой именно комплекс из двух сравниваемых меньше и характеризуется показанной на гистограмме долей.
РЕЗУЛЬТАТЫ
Гистограммы для перечисленных выше комплексов см. Юрский комплекс , Ундинодаинский комплекс, Тургинский комплекс, Исяньский комплекс, Андахудукский комплекс, Зазинский комплекс, Хурилтский комплекс.
На каждой гистограмме левый столбец соответствует комплексу, наиболее близкому к рассматриваемому. Вправо сходство убывает. Последовательность столбцов на гистограммах для родов и родов вместе с видами с разными весами может несколько меняться. Для восприятия и оценки всей совокупности результатов желательно свести их в более компактную форму.
Сходство глобальных фаун уменьшается при их удалении друг от друга во времени вследствие постоянного появления и вымирания таксонов. Это же может проявиться и при работе с региональными фаунами, но только как тенденция. Монотонное снижение сходства глобальных фаун в этом случае может искажаться в результате действия разнообразных факторов: экологических, тафономических, биогеографических (если изучаемый регион большой), ошибочной корреляции местных стратонов и местонахождений, неоднородности сборов в разных местонахождениях, неполноты изученности коллекций, различий в мнениях разных палеонтологов (? и др.).
На гистограммах рядом располагаются столбцы (комплексы), сходным образом удаленные от комплекса, с которым проводится сравнение. Можно использовать это обстоятельство для общей оценки всех результатов вместе.
Подсчитывалось число одноименных пар смежных столбцов на всех гистограммах. Например,
турга рядом с зазой 4,
юра рядом с андахудуком 5.
Сближение юрского и андахудукского комплексов обнаруживается на гистограммах для ундинодаинского, тургинского и исяньского комплексов. Но андахудукский комплекс характеризуется присутствием Hemeroscopus и заведомо моложе тургинского и исяньского компексов с Sinaeschnidia. Также он моложе ундинодаинского комплекса, более древнего, чем тургинский. На указанных гистограммах он оказался рядом с юрским, так как удален от этих комплексов сходно с юрским, но поскольку располагается от них в другую сторону на шкале времени этот и все подобные случаи следует исключить из подсчета, так что вторая строка примера в данном случае должна быть записана:
юра рядом с андахудуком 5-5=0.
В итоге выявляются следующие ряды (в скобках – число столбцов, смежных с выделенным полужирным шрифтом).
Юра → унда (11) → турга (10) → исянь (6) → андахудук (0) → заза (0) → хурилт (0)
Юра (11) ← унда → турга (10) → исянь (2) → андахудук (1) → заза (0) → хурилт (0)
Юра (10) ← унда (10) ← турга → исянь (9) → андахудук (3) → заза (2) → хурилт (0)
Унда (2) ← юра (6) ← турга (9) ← исянь → андахудук (8) → заза (7) → хурилт (3)
Юра (0) ← унда (1) ← турга (3) ← исянь (8) ← андахудук → заза (16) → хурилт (4)
Юра (0) ← унда (0) ← турга (2) ← исянь (7) ← андахудук (16) ← заза → хурилт (12)
Юра (0) ← унда (0) ← турга (0) ← исянь (3) ← андахудук (4) ← заза (12) ← хурилт
Порядок комплексов за одним исключением одинаков во всех рядах и соответствует их наиболее правдоподобной хронологической последовательности, как она реконструируется по всей совокупности данных. Юрский комплекс оказывается рядом с ундинодаинским в 11 случаях, тургинским в 10, исяньским в 6 и нигде рядом с андахудукским, зазинским или хурилтским. Исяньский комплекс наиболее близок тургинскому и андахудукскому и т. д.
Возможно, данная последовательность является слишком дробной. Не исключено объединение андахудукского комплекса с зазинским. Тем не менее, далее в качестве рабочей гипотезы принимается именно эта последовательность, так как в таком виде она более перспективна в качестве основы для дальнейших исследований.
ПОБОЧНЫЕ РЕЗУЛЬТАТЫ
Местонахождения, не учтенные выше из-за неочевидности их стратиграфического положения, можно попытаться датировать, используя полученную последовательность в качестве региональной хронологической шкалы (она может оказаться с перекрытиями или неполной, с лакунами). Высота столбцов на гистограммах в этом разделе равна числу баллов, группы из трех столбцов расположены в порядке принятой здесь хронологической шкалы, в каждой группе левый столбец – роды, средний столбец – роды и виды с весом 2, правый столбец – роды и виды с весом 5.
Улугейская свита (Хутулийн). Небольшой комплекс (8 родов, причем 2 – формальные роды жуков по надкрыльям). Состав мало диагностичен для определения возраста. Половина родов есть в юре. Присутствуют меловая Coptoclava longipoda, Cretobraconus, известный в Хотонте, и предположительно Astrocorethra, найденная в гурванэренской свите. По положению в разрезе (?) и косвенным соображениям об эволюции вулканизма свита должна относиться, по сообщению А.Г. Пономаренко, к самой нижней части мела. Скорее всего, возраст свиты тургинский.
По описанию разреза в книге С.М. Синицы (1993, стр. 157) слой с Coptoclava залегает в нескольких сантиметрах выше слоя с многочисленными изофлебиидными стрекозами, что считается характерным признаком отложений конца юры. Все же вряд ли на этом основании следует провести границу систем с небывалой точностью между коптоклавой и стрекозами, так как изофлебииды известны и в более молодых отложениях (в том числе в Байсе), а их род в Хутулийне не определен.
Кутинский горизонт . В забайкальских стратиграфических схемах кутинский горизонт относят к апту-альбу. Комплекс насекомых не моложе тургинского (менее вероятно, исяньского). Возможно, насекомые происходят из нижней части кутинского горизонта. Определение возраста как тургинского (подразумевается используемая здесь насекомная последовательность) относится только к отложениям, из которых собраны насекомые, но не кутинскому горизонту в целом
Семён
. Больше
всего зазинских элементов и много хурилтских. Cionocoleus не известен выше андахудукского горизонта. Неожиданным является присутствие характерного тургинского Ephemeropsis trisetalis (есть также в кутинских отложениях). Семен предположительно отнесен к андахудукскому
уровню, чтобы не расширять распространение E. trisetalis еще дальше вверх.
Черновские копи. Список родов (Clavineta, Siberiogenites, Trianguliperla, Mesoneta, Karanemoura, Mesobaetis, Stackelbergisca) выглядит юрским, но только два рода из семи (Karanemoura и Stackelbergisca) не найдены в мелу. Нет общих с другими комплексами видов.
По сообщению Д. Василенко, читинские геологи-съемщики относят отложения Черновских копей и Семёна к доронинской свите. Насекомые совершенно различны. Объединение этих местонахождений в составе одной свиты может быть оправданным только в случае исяньского возраста Черновских копей. Это мало вероятно, но не исключено полностью, так как в исяне много юрских родов (см. гистограмму на стр. 7) и в отдельных сравнительно небольших коллекциях исяньского возраста они могут оказаться преобладающими просто из-за случайности выборки.
Нижнецаганцабская подсвита
. Много юрских представителей. При учете видов на первый план выходит
сходство с тургинскими насекомыми, есть Sinaeschnidia.
Подсвита достаточно уверенно сопоставляется с тургинскими отложениями.
Хотонт . Небольшой комплекс с присутствием общих элементов с моготуинской, гурванэренской свитами и нижнецаганцабской подсвитой при значительном участии юрских компонент. Есть Manlaya, появляющаяся в турге, нижнецаганцабский вид Juleyrodes gilli и Siberiogenites cf. medius, похожий на нижнецаганцабского S. medius. Предположительно тургинский горизонт.
Дорготская свита. Всего 4 рода (Agrilium, Baleyopteryx, Chironomaptera, Siberioperla). Видов, известных в других комплексах нет. Все 4 рода есть в юре, Siberioperla также в ундинодаинском горизонте, а Baleyopteryx в Хотонте. Свита вряд ли моложе тургинского горизонта.
Верхнецаганцабская подсвита
. Небольшой
комплекс, сильно отличающийся от нижнецаганцабского из Хутел-Хары. Гистограмма не
дает отчетливой картины. При учете видов с большим весом она показывает
несколько большее сходство с андахудукским комплексом, но это определяется
присутствием только двух видов (Ephemeropsis melanurus и Notonecta xyphiale), причем
второй известен и в Исяне. Подсвита согласно
залегает на нижнецаганцабской (в Цаган-Цабе). Поэтому более вероятен исяньский
возраст, но с исянем общего только Chironomaptera gregaria и Notonecta xyphiale. Для точного заключения нужно найти Sinaeschnidia или Hemeroscopus.
Моготуинская свита . Гистограмма такая же неопределенная, как в предыдущем случае. Возраст может оцениваться в пределах от тургинского до андахудукского.
Гурванэренская свита. Сходная картина, по гистограмме трудно сделать достаточно правдоподобное заключение.
Последние три комплекса демонстрируют сложность датировки в случае небольшого количества данных. Объем этих проблемных комплексов ограничен имеющимися списками, но сравниваемые с ними основные фауны могут быть увеличены. Если принять полученную выше последовательность комплексов, то в таблице основных комплексов со столбцами, расположенными в порядке этой последовательности, можно заполнить пустые клетки между непустыми. Тогда состав основных фаун будет охарактеризован более полно и сравнение с ними может стать более информативным (см. “Сравнение с расширенными основными комплексами” для каждого из трех стратонов). Во всех трех случаях гистограммы несколько изменились, но неопределенность датировки устранить не удается. Присутствие Ephemeropsis melanurus в верхнецаганцабской подсвите и в моготуинской свите скорее всего исключает их тургинский возраст, о том же самом может свидетельствовать Notonecta xyphiale в гурванэренской свите, так что основной выбор относится к их исяньскому либо андахудукскому возрасту.
В качестве диагностического признака, позволяющего осуществить этот выбор, может быть использована доля (процент) юрских компонент в проблемных комплексах, иллюстрируемая следующей гистограммой:
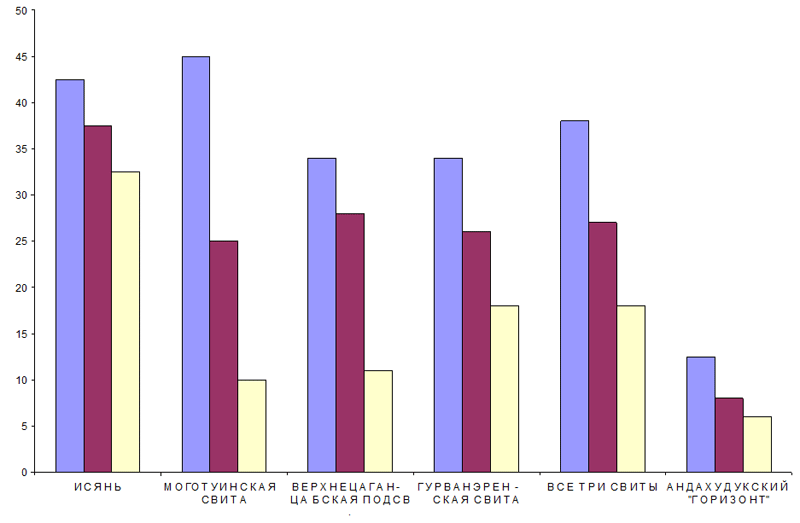
В каждой группе левый столбец показывает долю известных в юре родов среди родов комплекса, обозначенного в подписи у горизонтальной оси, средний и правый столбцы – то же самое для родов и видов с весом 2 и 5 соответственно. Так как число баллов в каждом из трех стратонов (особенно в моготуинский свите и верхнецаганцабской подсвите) невелико, 95-процентный доверительный интервал для доли сравним по величине с долей, показанной на гистограммах. Чтобы повысить достоверность доли в предпоследней группе из трех столбцов даны объединенные результаты по всем трем проблемным комплексам. В этой группе доверительный интервал 14.4 для родов, 11.0 для родов и видов с весом 2 и 8.1 для родов и видов с весом 5. Таким образом, роль юрских элементов резко снижается в андахудукское время, а проблемные комплексы по этому показателю близки к Исяню.
Отложения с Hemeroscopus
Хобур. Есть байсинский Diapherinus ornatipennis, известный также из андахудукского горизонта (Анда-Худук, Шин-Худук).
Эрдени-Ула (хухтыкская свита). Тоже байсинско-андахудукская (Цогт-Обо) Hydrophilopsia baissensis.
Баян-Дурах (предположительно андахудукская свита). Найдены Ephemeropsis melanurus и ?Carabites cf. caraboides (ещё в кутинском горизонте, Борзя-II).
Западная Эрдене-Ула (ундурухинская свита). Отнесена в эту группу местонахождений условно. Здесь нет Hemeroscopus, есть большой комплекс домиков (23 вида) и жук Brochocoleus planus, найденный также в Бон-Цагане (род известен, начиная с Исяня).
Первые три местонахождения андахудукского или байсинского возраста, последнее вряд ли древнее андахудукского.
Китай
В сводном разрезе провинции Ляонин и прилегающих к ней частей Внутренней Монголии и провинции Хебей сверху вниз различаются следующие свиты (Ren and oth., 1995) (полужирным шрифтом выделены свиты с насекомыми, прямым полужирным – с очень крупными местонахождениями
Лушанфен (Lushangfen = Fuxin)
Шахай (Shahai)
Юфотан (Jiufotang)
Исянь (Yixian)
несогласие
Жаньяку (Zhangjiakou)
Хучен (Houcheng = Tuchengzi)
Тяоишан (Tiaojishan) с туфогенно-осадочной пачкой Даохугоу (Daohugou) внизу
несогласие
Юлоншан (Jiulongshan) и Хайфангоу (Haifanggou)
Ранее пачка Даохугоу и формации Юлоншан и Хайфангоу считались относящимися к одному
стратиграфическому уровню (так в указанной выше монографии). Современные представления о соотношениях
формации Юлоншан и пачки Даохугоу иллюстрируются следующим рисунком из Liu et al., 2006:
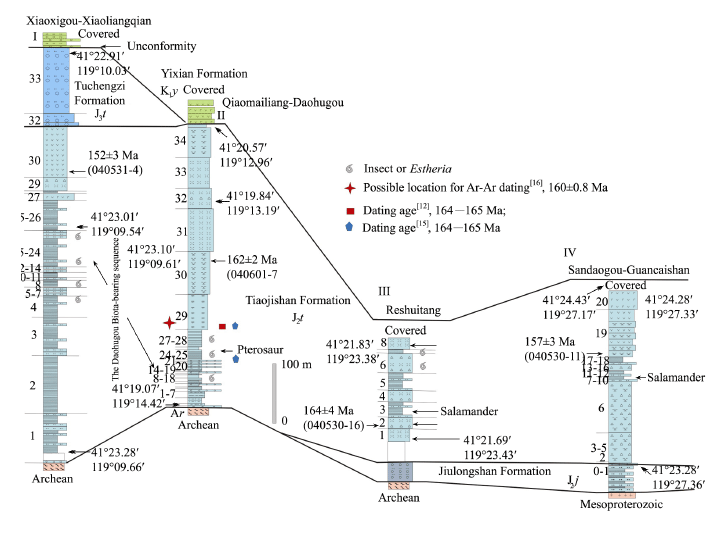
Стратиграфическое положение формации Хайфангоу такое же, как у
формации Юлоншан, обе формации считаются синхронными.
Радиоизотопный возраст пачки Даохугоу около 164 млн. лет (келловей). Zhang Junfeng (2014) составил таблицу с общими родами насекомых из Каратау, пачки
Даохугоу и формаций Юлоншан и Хайфангоу. В ней обращает на себя внимание 31
каратаусский род в Даохугоу и только три из них в формации Юлоншань. В табличке
почему-то не приведены роды, общие для Юлоншаня и Каратау и не найденные в
пачке Даохугоу, хотя такие роды, конечно, должны быть.
По данным PaleoDB и EDNA в Даохугоу всего 260 родов, 48 родов, общих с другими местонахождениями,
из них с Каратау – 24 (50%), с Юлоншанем – 14 (29%). В формации Юлоншань –
всего 118 родов, 33 рода, общих с другими местонахождениями, из них с Каратау –
13 (39%), с Даохугоу те же 14 (42%).
Очевидно, даохугоусский комплекс моложе юлоншаньского, но трудно
определить насколько моложе. Для основной работы по эволюции разнообразия
насекомых принят келловейский возраст Даохугоу и батский Юлоншаня и Хайфангоу.
Хучен. Бедный комплекс всего с 9 родами, но только три из них известны в других местах (Rhipidoblattina, Protorthophlebia, Sogdoblatta). Все три есть в юре, первый, кроме того, в исяне и гурванэренской свите. Данные по абсолютному возрасту противоречивы – 135 и 152 млн. лет.
Юфотан . По возрасту свита должна быть как Исянь.
Шахай Свита согласно залегает на Jiufotang fm. с более богатым комплексом исяньского состава и согласно перекрывается Lushangfen fm. с Hemeroscopus baissicus. Из 5 родов в составе комплекса (Holcorobeus (Geotrupoides), Sinaeschnidia, Rhipidoblattina, Aboilus, Chengdecupes) три последние присутствуют в юре. Подрод Geotrupoides есть в Байсе, а также в нижнецаганцабских и гурванэренских отложениях Монголии. Присутствует исяньско-лайянская Sinaedischnia heishankowensis. По возрасту свита должна быть отнесена к исяню.
Лушанфен
. По общему составу
комплекс ближе всего к исяньскому, но появляются андахудукские, зазинские и
хурилтские представители. Только для андахудукского горизонта известно
сочетание первых Hemeroscopus и Cionocoleus, не найденного в заведомо более молодых отложениях.
С учетом согласного залегания на формации Шахай и присутствия Hemeroscopus baissicus возраст андахудукский.
Лайян. Известные насекомые формации Лайян (Laiyang) происходят из провинции Шаньдун
(в 300-
ВИДЫ
Все данные о видах, связывающих разные комплексы, сведены в таблицу, в которой кроме основных комплексов есть несколько дополнительных. Для этих комплексов использованы следующие обозначения: Kuti - кутинский, LTsag - нижнецаганцабский, UTsag - верхнецаганцабский, Mogo - моготуинский, Gurv - гурванэренский, как Khot и Seme обозначены комплексы из местонахождений Хотонт и Семен соответственно. На таблице видно следующее:
1. Много общих видов в ундинодаинском и тургинском комплексах (нижняя левая часть таблицы). Скорее всего, между ними нет существенного временнóго зазора.
2. Кутинский комплекс небольшой, не отделяется от тургинского.
3. Хорошо связанную группу образуют андахудукский, зазинский и хурилтский комплексы вместе с насекомыми Семена.
4. Нижняя правая и верхняя левая части таблицы практически пустые. В тургинском комплексе общие с предыдущей группой только Ephemeropsis trisetalis (Семен) и Eoanthocoris cf. cretaceous (Бон-Цаган).
5. Имеется довольно рыхлый кластер (выделен светло-зеленым прямоугольничком), в котором объединяются “проблемные” моготуинский, гурванэренский и верхнецаганцабский комплексы. В нем обычно имеется только по 2 вида, общих для этих комплексов попарно. Такие же слабые связи обнаруживаются с исянем (если включить сюда лайянскую Chironomaptera vesca), тургой и андахудукско-хурилтским комплексом (если не учитывать широко распространенные Ephemeropsis melanurus и Notonecta xyphiale). Не вполне отчетливая обособленность этих трех комплексов по видам хорошо согласуется с неопределенностью их возраста по гистограммам.
6. У Исяня нет хорошей характеристики по пригодным для сопоставлений видам. Имеются только Notonecta xyphiale, широко встречающаяся в слоях с Hemeroscopus, Mesypochrysa cf. chrysopoides, описанная из Бон-Цагана, зазинская Parahagla sibirica и два вида Chironomaptera из монгольских проблемных комплексов. На таблице исяньские виды ближе к правой верхней части, общего с тургой только два сомнительных вида хирономаптер.
БИОСТРАТИГРАФИЯ
Для каждого горизонта можно привести список родов и видов, найденных только в нем и потенциально пригодных для корреляции отложений в пределах горизонта и датирования новых коллекций (в принимаемой здесь региональной шкале). Дополнительные возможности могут дать совместные находки родов и видов, встреченных в отложениях определенного горизонта в первый и в последний раз. Этот способ будет работать, если последовательность горизонтов надежно установлена и их фауна достаточно изучена. Многие такие таксоны или пары таксонов известны только в одном местонахождении или в одной свите. Роды и виды, известные только в одном горизонте, часто представлены немногими или единственным экземпляром. Стратиграфическое значение таких родов и видов или пар таксонов фактически не выяснено.
Основной интерес представляют роды, виды и пары таксонов, найденные в одном горизонте в разных свитах, демонстрирующие реальную возможность их использования для стратиграфии. Ниже приводится обзор таких таксонов по горизонтам.
Ундинодаинский горизонт
Только в ундинодаинском горизонте в разных свитах найдены следующие роды и виды:
Furvoneta lata - глушковская и укурейская свиты,
Karatoma raptor - глушковская и укурейская свиты,
Mesocorethra
Mesohelophorus elongatus - глушковская и укурейская свиты,
Necrotaulius minutissimus - глушковская и укурейская свиты,
Oryctochlus mirificus - глушковская и укурейская свиты,
Unda pachycephala - глушковская и укурейская свиты,
Bryanka - глушковская и укурейская свиты (B. lepida, B. venusta, B. venustula - глушковская свита, B. antis - укурейская свита),
Dahurium - глушковская и укурейская свиты (D. draco - глушковская свита, D. cf. draco - укурейская свита),
Sciaromima - глушковская и укурейская свиты (S. minor – глушковская свита, S. major – укурейская свита),
Sibelcana - глушковская и укурейская свиты (S. rossica – глушковская свита, S. transbaicalica – укурейская свита),
Sinitsia – глушковская, укурейская и тергенская свиты,
Flexoperla - укурейская и чалунихинская свиты (F. recusata - укурейская свита, F. flexuosa - чалунихинская свита);
И в глушковской и в укурейской свитах имеется 15
одинаковых пар родов или видов, диагностичных для ундинодаинского горизонта (первые
Mesohelophorus, Furvoneta undina, Liadytes longus, Mesosperchus tarsalis, Proameletus caudatus и последние Karatoma, Necrotaulius, Pleciomima).
Стрекозы рода Xeta (укурейская свита) кроме Забайкалья есть в уланэрегской свите в Монголии (Хоутийн-Хотгор); возможно свита одновозрастна с ундинодаинскими отложениями.
Приведенные данные определенно подтверждают правильность корреляции глушковской и укурейской свит и менее уверенно их одновозрастность тергенской и чалунихинской свитам. Насекомые относимой к ундинодаинскому горизонту букукунской свиты не включают родов, видов или пар первых и последних находок, стратиграфическое значение которых может считаться подтвержденным.
Тургинский горизонт
Руководящий комплекс тургинского горизонта составляют в основном 6 видов клопов. Сопоставление свит по насекомым иллюстрируется следующей таблицей (знаком + обозначены пары первых (до знака +) и последних (после +) находок.
|
|
Turga |
Gidara |
Daya |
Byankino |
UstKara |
|
Aphanocoris femoratus |
X |
X |
|
|
|
|
Aphanocoris porrectus |
X |
|
X |
|
|
|
Corienta transbaicalica |
X |
X |
|
|
|
|
?Holcorobeus incertus |
X |
X |
|
|
|
|
Positocoris intermedius |
X |
|
X |
|
|
|
Positocoris sibiricus |
X |
X |
|
|
|
|
Takshania implicata, |
X |
X |
|
|
|
|
Aphanocoris turgensis + Proameletus |
X |
X |
|
X |
|
|
Coptoclava + Mesosperchus |
X |
|
|
X |
X |
|
Coptoclava + Proameletus |
X |
X |
|
X |
X |
|
?Coptoclavella + Liadytes longus |
X |
|
X |
|
|
|
Corienta + Proameletus |
X |
X |
|
|
|
|
Ephemeropsis + Proameletus |
X |
X |
|
|
|
|
Manlaya + Proameletus |
X |
X |
|
|
|
|
Mesogyrus striatus
+ Liadytes longus |
X |
|
X |
|
|
|
Mesogyrus striatus
+ Mesosperchus |
X |
|
|
X |
|
|
Mesogyrus striatus
+ Proameletus |
X |
|
X |
X |
|
По совокупности палеоэнтомологических данных объединение всех перечисленных свит в единый горизонт является вполне обоснованным. Годымбойская и даинская свита в местных стратиграфических схемах относятся к верхней части тургинского горизонта. По насекомым они не отличаются от других тургинских свит, но коллекции сравнительно малы. В устькарской свите Coptoclava найдена в Полосатике, а Mesosperchus и Proameletus в Усть-Каре, так что датировка по сочетанию родов зависит от корректности объединения этих местонахождений в составе одной свиты. Годымбойская свита не содержит диагностических для горизонта насекомых.
Исяньский “горизонт”
На исяньском уровне основной материал происходит их одного местонахождения в формации Исянь. Другие местонахождения в этой формации дают мало дополнений. Поэтому здесь трудно выделить какие-либо роды и виды, потенциально пригодные для корреляции отложений. Как уже упоминалось, в исяньском комплексе присутствует около 40% родов, известных в юре (см. гистограмму на стр. 6), так что не исключен юрский облик некоторых локальныъ фаун исяньского возраста.
Андахудукский горизонт
Только в андахудукской и шарынгольской свитах есть Zygadenia caudatus, не найденная нигде больше. Андахудукскую и шинхудукскую свиты объединяет присутствие последних Zygadenia и первых Diapherinus ornatipennis, Hemeroscopus baissicus или Notocupes cf. elegans. Нет данных для непосредственного сопоставления шинхудукской и шарынгольской свит. Тэвшская свита и уланаргалантские отложения района Бон-Цагана. по присутствию Hemeroscopus baissicus вместе с Ephemeropsis и стратиграфическому положению ниже хурилтской свиты в районе Бон-Цагана и шартологойской в Шар-Тологое могут относиться к андахудукскому или зазинскому уровню.
Скудные возможности для корреляции по насекомым в андахудукском горизонте прямо связаны с сравнительно малым материалом из этих отложений. Так, в тургинском горизонте имеется 117 родов или видов, найденных только в нем, 30 первых находок и 23 последних, тогда как в андахудукском соответственно только 20, 12 и 10. Примерно так же соотносятся и полные списки комплексов.
Зазинская свита и хурилтский горизонт
Зазинский комплекс известен только из Байсы (в единичных случаях также из Романовки), так что выделить стратиграфически полезные таксоны невозможно. Для хурилтского горизонта данные о последних находках не могут быть использованы, так как более молодые энтомофауны в центральноазиатских нижнемеловых отложениях не найдены. В хурилтской и шартологойской свитах найдены отсутствующие в других горизонтах Cretanaides, Lithoscarabaeus, Metabuprestium dundulense, Valditermes acutipennis.
Замечания
1. Приведенные выше насекомные маркеры (таксоны и пары таксонов) позволяют подтвердить корреляцию некоторых свит ундинодаинского, тургинского и андахудукского горизонтов и демонстрируют возможность их сопоставления по насекомым. Но маркеры выявлены с использованием этой же корреляции, изначально принятой для последующего обобщения данных. В результате имеется логический круг, и они не добавляют ничего нового для самого сопоставления свит.
Выход может быть в переходе к работе на уровне свит. Однако, на примере андахудукского горизонта, а также моготуинской, гурванэренской свит и верхнецаганцабской подсвиты хорошо видно резкое снижение стратиграфического потенциала насекомых при уменьшении объема коллекций. Это связано с высоким богатством энтомофауны, так что вероятность находок одного таксона в двух коллекциях сильно снижается для небольших коллекций (богатство водных насекомых значительно меньше и находки одних и тех же родов и видов в разных коллекциях намного вероятнее). Видимо, с этим связана неудачная попытка анализа комплексов по свитам, сделанная в начале данного исследования. По той же причине еще более затруднен переход к непротиворечивой стандартной биостратиграфической методике, основанной на послойном изучении коллекций. Кроме того, ее применение чрезвычайно ограничено фрагментарностью материала: в тех немногих случаях, когда ясна последовательность свит, она как правило не охарактеризована представительными комплексами насекомых.
2. Стратиграфическое значение маркеров состоит, главным образом, в их потенциальной пригодности для датировки новых коллекций. Однако, пример проблемных комплексов показывает ограниченные возможности реального применения маркеров. Это связано все с той же фрагментарной изученностью чрезвычайно богатых древних энтомофаун.
3. По тем же причинам в настоящее время затруднительно получение
обоснованных датировок при работе с отдельными группами насекомых.
4. Польза маркеров состоит, главным образом, в возможности дать обоснованные списки руководящих комплексов хотя бы для Восточного Забайкалья, то есть получить результаты в стандартной для биостратиграфии форме.
5. В коллекциях из хурилской и шартологойской свит отсутствуют поденки. Ephemeropsis вероятно вымер
к этому времени, но его нет в Исяне, Хутел-Харе и гурванэренской свите, хотя он
несомненно существовал во время накопления соответствующих отложений. Ephemeropsis trisetalis в Семене не древнее андахудукского уровня. E. melanurus есть в Анда-Худуке, верхнецаганцабской подсвите и моготуинской свите. Распространение этих двух видов перекрывается по крайней мере в андахудукское время, и, возможно, в исяне.
6. Формальный род Chironomaptera объединяет изолированные крылья хаоборид, которые могут сильно отличаться по личинкам (Калугина, 1993). Это не исключает возможности ограниченного использования видов Chironomaptera в биостратиграфии, но предположительные определения видов этого рода для стратиграфических выводов мало пригодны. Последние Chironomaptera указываются в Исяне. Chaoboridae присутствуют и выше, до хурилта, но либо не изучены, либо только куколки, как в Байсе. Интерес представляет более точное определение хотя бы бонцаганских хаоборид.
7. Основные монгольские местонахождения, Исянь, Байса и Семен моложе восточнозабайкальского тургинского горизонта. В имеющихся монгольских коллекциях тургинский уровень представлен очень слабо, только в Хутулийне, Хутэл-Харе и Хотонте, а в Китае не известен. Ундинодаинскому горизонту, возможно, соответствует уланэрегская свита (Хоутийн-Хотгор). Не исключена биогеографическая специфика Восточного Забайкалья в конце юры и раннем мелу, но данных недостаточно. В этом случае, возможно, монгольская фауна пришла в Забайкалье в андахудукское время (Семен). Большой интерес представляет изучение верхней части кутинского горизонта и отложений гусиноозерской зоны Забайкалья.
ДАТИРОВКИ
Рубеж юры и мела предположительно совпадает с появлением массовых
захоронений Coptoclava и Ephemeropsis и находками в тургинских отложениях ряда
других родов и видов (см. таблицу основных комплексов).
Рубеж Sinaeschnidia/Hemeroscopus датируется
радиоизотопным возрастом Исяня как граница баррема и апта. Эта датировка
получена аргон-аргоновым методом по санидинам из
прослоя вулканического пепла в отложениях с исяньской фауной (Swisher and oth., 1999). Позднее она была
подтверждена тем же методом в контрольном исследовании авторитетной Ламонтской
обсерватории (Chang and oth.,
2009). Имеется также независимая совпадающая датировка более надежным уран-свинцовом методом по цирконам (Renne and oth., 2010). В настоящее
время эта датировка является общепризнанной среди специалистов по
радиоизотопному датированию (Gradstein and oth.,
2012).
Тургинский и сменяющий его исяньский комплексы существовали в течение берриаса-баррема.
Фауна исяньского типа широко распространена и повидимому существовала длительное время. За неимением данных, определяющих положение ее нижней границы, не остается ничего, кроме произвольного деления всего интервала пополам, то есть турга – берриас-валанжин, исянь – готерив-баррем.
Андахудук и заза попадают в апт.
Хурилтский комплекс хорошо бы
отправить в нижний альб. Это мнение не основано на
каких-либо данных, однако кажется очень странным слишком малое число альбских
каменных коллекций. Не результат ли это неточной датировки каких-то комплексов ? Симптоматичным может быть присутствие только в Бон-Цагане
сепульки Ghilarella, описанной из среднеальбской еманринской
свиты Хетаны.
Дополнительные сведения о датировках в принципе могут быть получены по
другим группам ископаемых с учетом более широких корреляций. Например, имеется
китайская статья (Sha et al., 2012) по двустворкам с корреляцией Исяня
на восток до Японии, где есть морской мел с аммоноидеями. В этой работе формация
отнесена к готериву-баррему. В данном обобщении сведения по другим группам не
использовались.
КОРРЕЛЯЦИЯ
Результаты проведенного обобщения суммированы в следующей таблице:
|
Возраст |
Уровень (“горизонт”) |
Забайкалье |
Монголия |
Китай |
|
Апт (? нижний альб) |
Бонцаганский |
|
хурилтская свита (Бон-Цаган, Хурилт, Холботу-гол), Шар-тологой |
|
|
Апт |
Зазинский |
Байса, Романовка |
|
|
|
Апт |
Андахудукский |
Семен |
Анда-худук, Шин-худук, Шарын-гол |
формация Лушанфен |
|
Готерив-баррем |
Исяньский |
|
верхнецаганцабская подсвита (Цаган-цаб, Ринчин-бумба, Цаган-субурга, Ундур-шиль), моготуинская свита (Гер-обо, Гер-обо-шине, Манлай, Толь, Улан-Тологой, Хабтагай-ула), гурванэренская свита (Гурван-эрен-нуру, Хух-морт, Мянгад, Алтан-теели, Боро-нуру) |
формации Исянь, Юфотан, Шахай, Лайян |
|
Берриас-валанжин |
Тургинский |
Тургинский и нижняя часть кутинского горизонта |
Хутэл-хара, Хутулийн (только верхняя часть разреза ?), Хотонт |
|
|
Титон |
Ундинодаинский |
Ундинодаинский горизонт |
Хоутийн-хотгор |
|
ЗАКЛЮЧЕНИЕ
Изложенные выше результаты следует считать предварительными. Для
получения более обоснованной последовательности комплексов необходим полный
учет всего доступного и хорошо ревизованного фактического материала. Все же
полученная основная последовательность (см. "Результаты") выглядит достаточно
убедительной и, можно надеяться, сохранится. Она может измениться, если в
раннем мелу существовали серьезные биогеографические барьеры, разделявшие
рассмотренные здесь регионы. Этот вопрос не имеет решения, пока нет хорошо
охарактеризованной последовательности комплексов по каждому региону в
отдельности.
В связи с очень высоким разнообразием насекомых приемлемые результаты
удается получить при работе с обобщенными комплексами, составленными на основе
уже имеющейся корреляции отложений. Самостоятельное стратиграфическое значение насекомых поэтому продолжает оставаться ограниченным, но в
некоторых указанных выше случаях оно вполне очевидно (см. раздел “Биостратиграфия”).
Примененные в этой работе способы анализа данных могут быть полезны и в
дальнейших исследованиях.
Автор благодарен всем сотрудникам Лаборатории артропод, неизменно
консультировавшим его по всем возникавшим вопросам.
ЛИТЕРАТУРА
Атлас фауны и флоры палеозоя-мезезоя Забайкалья (под ред. А.Н. Олейникова).
Новосибирск: Наука, 2002. 713 с.
Дмитриев В.Ю., Аристов Д.С., Башкуев
А.С., Василенко Д.В., Вршанский П., Горохов А.В., Лукашевич Е.Д., Мостовский М.Б.,
Пономаренко А.Г., Попов Ю.А., Расницын А.П., Синиченкова Н.Д., Сукачева И.Д., Храмов
А.В., Шмаков А.С. Разнообразие насекомых от карбона до современности. Палеонтологический журнал. 2017 (в печати)
Калугина Н.С. Хаобориды и комары-звонцы из мезозоя Восточного Забацкалья (Diptera, Chaoboridae, Chironomidae) // Мезозойские насекомые и остракоды Азии. М: Наука, 1993. С. 117-139 (Тр. Палеонтол. ин-та АН СССР. Т. 252)
Калугина Н.С., Ковалев В.Г. Двукрылые насекомые юры Сибири. М.: Наука. 1985. 198 с.
Мезозойские насекомые и остракоды
Азии // Тр. Палеонтол. ин-та АН СССР. Т.
Позднемезозойские насекомые Восточного
Забайкалья // Тр. Палеонтологического института АН СССР. 1990. Т. 239. 223 с.
Синица С.М. Юра и нижний мел
Центральной Монголии // Тр. Совместной Советско-Монгольской палеонтологической
экспедиции. 1993. Вып. 42. 239
с.
Юрские насекомые Сибири и Монголии (ред. А.П. Расницын). М.: Наука, 1985. 193 с. (Тр. Палеонтол. ин-та АН СССР. Т. 211)
Chang
Su-chin, Zhang Haichun, Renne P.R., Fang Yan. High-precision
40Ar/39Ar age for the Jehol biota.
Palaeogeography, Palaeoclimatology, Palaeoecology. 2009. 280. P. 94-104.
Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg
G.M. (eds.) The geologic time scale 2012.
Liu Yongqing, Liu
Yanxue, Jl Shu’an, Yang Zhiqing. U-Pb zircon age for the Daohugou biota at
Ningcheng of Inner Mongolia and comments on relates issues. Chinese Science Bulletin, 2006. V.
51, № 21. Р. 2634-2644.
Ren D.Z., Guo L., Lu
L., Ji S., Han Y. Faunae and stratugraphy of Jurassic – Cretaceous in Beijing
and the adjacent areas.
Renne, P.R., Mundil, M., Balco, G.,
Min, K., Ludwig, K.R., 2010. Joint determination of 40K decay constants and
40Ar*/40K for the
Sha Jin-geng, Pan Yan-hong, Wang Ya-qiong,
Zhang Xiao-lin, Rao Xin. Non-marine and marine
stratigraphic correlation of Early Cretaceous deposits in NE China, SE Korea
and
Swisher III, C.C., Wang, Y.Q., Wang, X.L., Xu,
X., Wang, Y., 1999. Cretaceous age for the feathered
dinosaurs of
Zhang Junfeng.
Archisargoid flies (Diptera, Brachycera, Archisargidae and Kovalevisargidae)
from Jurassic Daohugou biota of